ヘキサラファンはKeap1とGsk-3βを同時に阻害する ― Nrf2を「二重経路」で活性化する最新研究を解説
本わさびに含まれるヘキサラファン(6-MSITC)は、これまで主に Nrf2を活性化する天然成分 として注目されてきました。ところが2025年に発表された最新研究は、ヘキサラファンの働きが「Nrf2のブレーキを1つ外す」だけでは説明しきれないことを示しました。1
今回取り上げるのは、García-Yagüeらが Free Radical Biology and Medicine に発表した論文です。本研究が示したのは、ヘキサラファンが従来知られていた Keap1(キープワン)のブレーキ解除 に加えて、Gsk-3β (ジーエスケースリーベータ)という別の分解経路も直接抑える可能性 です。つまりNrf2を守るしくみが「1本」ではなく「2本」あるかもしれない、ということです。加齢や慢性疾患で低下しやすい生体防御の理解を一段深める知見として報告されています。1
この記事で分かること
- ヘキサラファンが Keap1依存のNrf2活性化だけでは説明できない作用 を示したこと
- その追加経路として、Gsk-3β阻害によるNrf2分解の抑制 が示唆されたこと
- ブロッコリー由来のスルフォラファンには見られにくい特徴で、分子の構造差(炭素鎖の長さ) が関わる可能性があること
- 加齢や慢性疾患で Nrf2の低下とGsk-3βの亢進が重なる病態 を考えるうえで、意味のある示唆を持つこと
背景:Nrf2は「2方向」から分解されている
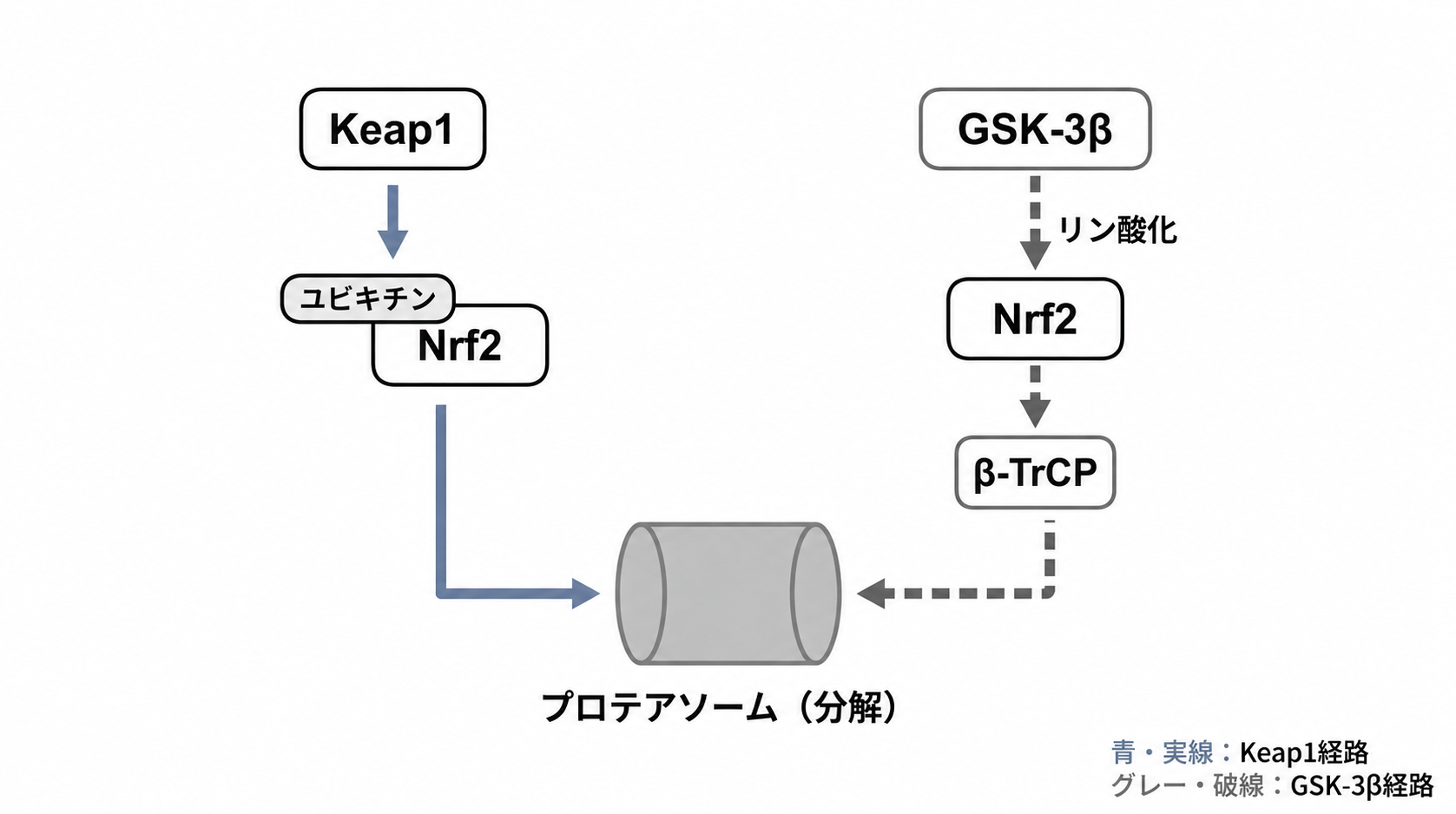
Nrf2は、必要なときだけ働けるように、ふだんは細胞の中で分解されています。その分解を担う代表的なしくみが、Keap1経路です(Nrf2の基本的な働きはこちらの記事で解説しています)。
Keap1はNrf2をつかまえ、ユビキチンという「処分の目印」を付けます。目印が付いたNrf2は、プロテアソームという分解装置へ運ばれます。Keap1は、Nrf2が増えすぎないようにするブレーキ役と考えると分かりやすいでしょう。3
ただし、Nrf2を分解する道筋はこれだけではありません。もうひとつ、Gsk-3β/β-TrCP経路と呼ばれるルートがあります。
このルートでは、まずGsk-3βがNrf2をリン酸化します。するとβ-TrCPがその変化を見つけ、Nrf2を分解装置へ送ります。Keap1経路とは別の方法で、Nrf2に「処分の合図」を出しているのです。2
 用語解説
用語解説- ユビキチン
- 不要になったタンパク質に付く「処分の目印」。
- プロテアソーム
- 目印の付いたタンパク質を分解する、細胞内の処理装置。
- リン酸化
- タンパク質にリン酸を付け、その働きや分解されやすさを変える反応。
- β-TrCP
- リン酸化されたNrf2を見つけ、分解へと導くタンパク質。
この2つ目のルートが注目される理由は、加齢やアルツハイマー型認知症、2型糖尿病などでは、Gsk-3βの働きが強くなると報告されているためです。つまり、こうした状態ではKeap1に加えてGsk-3βからも分解が進み、Nrf2が減りやすくなる可能性があります。3 4
たとえでいうと…
Nrf2は、いざというとき細胞を守る「防御チームのリーダー」です。ところが普段は、見張り役が2人います。
- Keap1:その場でリーダーに“処分タグ”を貼る係
- Gsk-3β:別の付箋を付け、運び屋にリーダーを連行させる係
ヘキサラファンは、この2人の見張り役のうち片方だけでなく、両方に働きかける可能性が出てきたのです。
この研究の問い:ヘキサラファンはKeap1以外にも働くのか
ヘキサラファンは、スルフォラファンと同様にKeap1を介したNrf2活性化が知られている成分です。1
ただし著者らが注目したのは、「ヘキサラファンの作用が本当にKeap1だけで説明できるのか」 という点です。過去には、6-MSITCが試験管レベルでGsk-3βを抑える可能性も報告されていましたが、細胞の中でNrf2の安定化につながるかどうかは十分に示されていませんでした。5
今回の論文は、この未解決の問いに対して、
- Keap1がない細胞でも作用するのか
- 間接的な経路ではなく、Gsk-3βそのものに働くのか
- スルフォラファンとの違いはどこにあるのか
を順に検証した研究です。1
研究でわかったこと
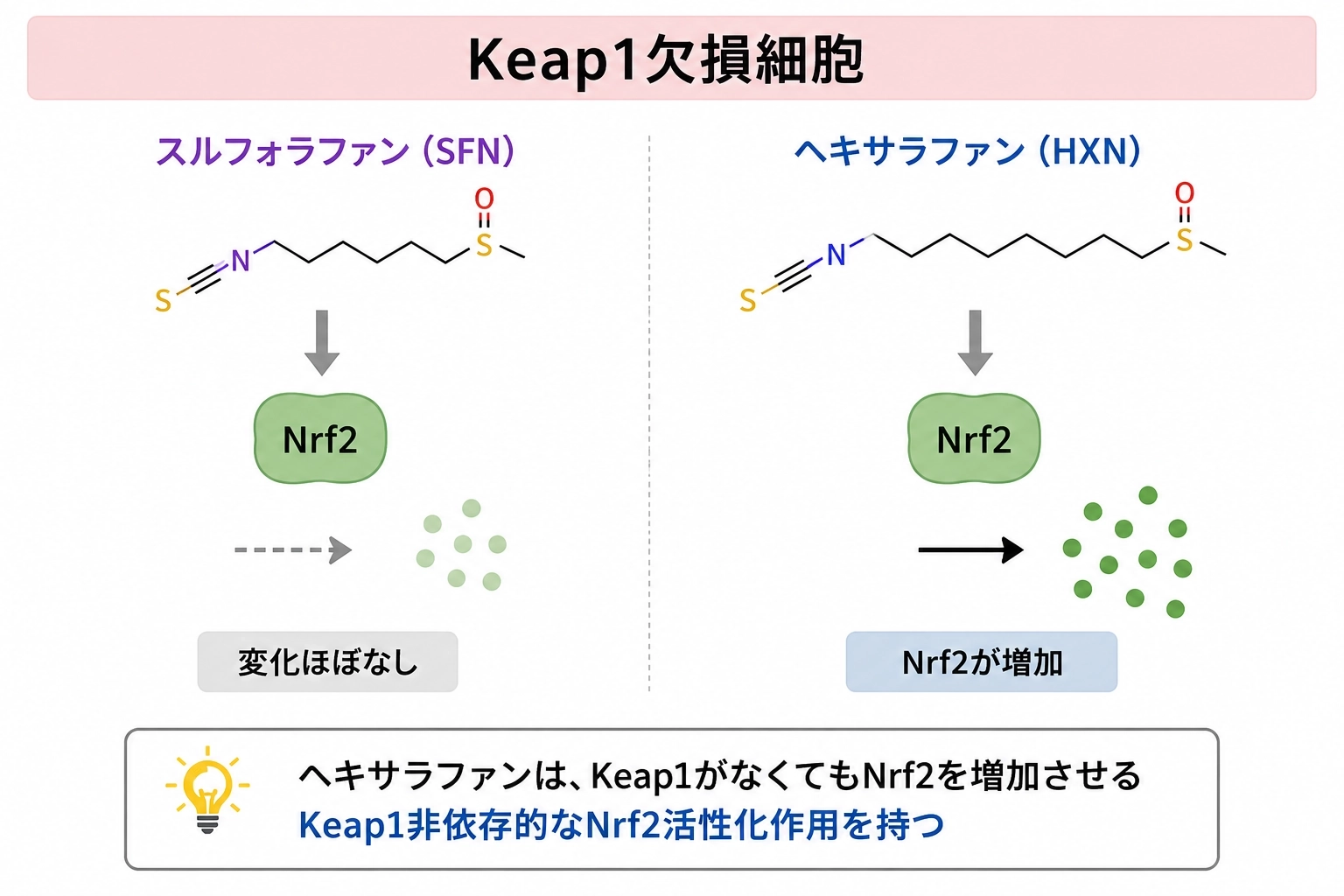
1. Keap1がなくてもNrf2が増えた
最初のポイントは、Keap1欠損細胞※でもヘキサラファンがNrf2を増やした ことです。Keap1がある通常の細胞では、ヘキサラファンもスルフォラファンもNrf2とその下流の防御タンパク質を増やしました。ところがKeap1を欠く細胞では、スルフォラファンの効果がほぼ消えたのに対し、ヘキサラファンはなおNrf2の増加を示しました。1
用語解説- Keap1欠損細胞
- Keap1を人工的に失わせた細胞。作用がKeap1に依存するかどうかを確かめる実験で使われる。
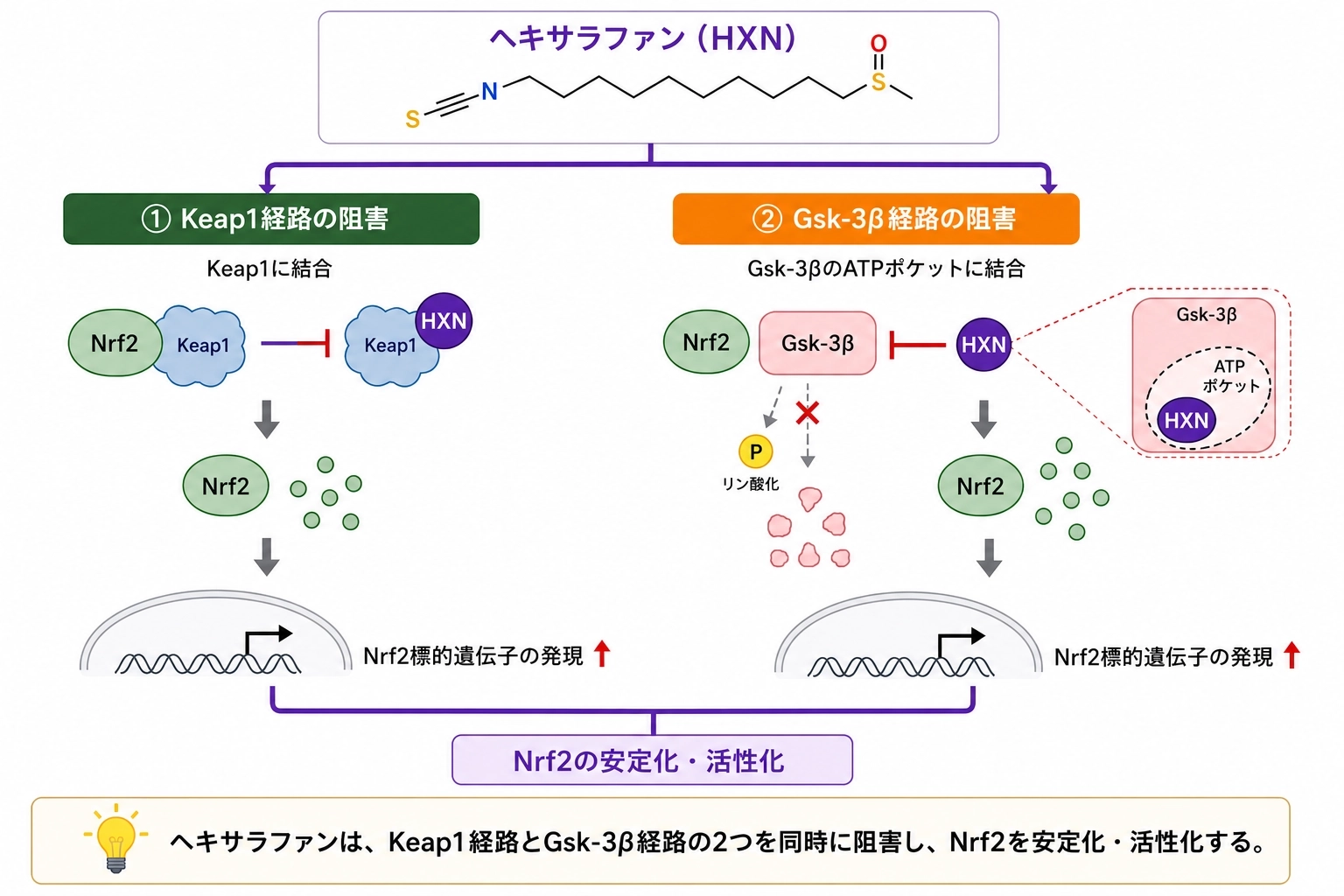
これは、ヘキサラファンには Keap1に依存しない作用成分が残っている ことを意味します。「Keap1を外す成分」という従来の理解だけでは不十分で、別の作用点がある可能性が示されました。
2. 代表的な間接経路は動かさなかった
Nrf2を高める物質の中には、Erk、p38Mapk、Jnk、Pten/Pi3k/Akt といった細胞内の経路を介して、間接的にGsk-3βを抑えるものがあります。ところが今回、ヘキサラファンは これらの代表的な経路に大きな変化を与えないまま Nrf2を増やしました。1
この結果は、「別の経路を回り道してGsk-3βを止めた」のではなく、Gsk-3β本体により近い形で作用している 可能性を後押しします。
3. Gsk-3β阻害でNrf2分解を止めた
研究の核心はここです。著者らは、ヘキサラファンが Gsk-3βそのものを抑えることでNrf2の分解を止める という仮説を、複数の実験で検証しました。1
主な結果は次のとおりです。
- Gsk-3βを活性化してNrf2を減らした条件でも、ヘキサラファンを加えるとNrf2の減少が抑えられた
- Nrf2に付くユビキチン化※の程度が、ヘキサラファンの濃度に応じて低下した
- その挙動は、既知のGsk-3β阻害剤SB216763とよく似ていた
- ヘキサラファンはNrf2のリン酸化を抑え、Gsk-3βとの結合も弱めた
用語解説- ユビキチン化
- タンパク質にユビキチンが付き、分解の対象として認識されやすくなる現象。
つまりヘキサラファンは、Nrf2に“分解の付箋”が貼られる前の段階を止めていた と考えられます。1
Keap1経路は「分解タグを貼る流れ」を止めるルート、Gsk-3β経路は「別の付箋を付けて分解へ回す流れ」を止めるルートです。今回の論文は、ヘキサラファンが後者にも働く可能性を、かなり具体的に示しました。
4. Nrf2のリン酸化を直接抑えた
さらに試験管内キナーゼアッセイ※では、ヘキサラファンが Gsk-3βによるNrf2のリン酸化そのものを抑える ことが確認されました。1
用語解説- キナーゼアッセイ
- リン酸化を担う酵素(キナーゼ)の働きを、試験管内で直接測定する実験。
これは「Gsk-3βに関連する周辺の経路を触った」のではなく、Gsk-3βの働きそのものを抑えている ことを、より直接的に支持する結果です。
5. 分子シミュレーションでもGsk-3βとの結合が支持された
コンピュータシミュレーション(分子ドッキング・分子動力学)では、ヘキサラファンがGsk-3βの 反応に関わるポケット(触媒溝)に比較的安定に収まり、既存の阻害剤に似た結合様式を示すと予測されました。1
著者らは、この結果からヘキサラファンが ATPポケット※付近を占有することでGsk-3βの活性を抑えている可能性 を考察しています。
用語解説- ATPポケット
- 酵素が働くためのエネルギー分子ATPが結合する部位。ここを別の分子が占有すると、酵素の活性が落ちることがある。
6. スルフォラファンとの違いは「炭素鎖の長さ」の可能性
ヘキサラファンとスルフォラファンはよく似た構造ですが、ヘキサラファンのほうが 炭素鎖が2つ長い のが特徴です。著者らは、この違いがGsk-3βのポケットへの結合の安定性に関わる可能性を示唆しています。1
つまり、似た仲間の成分でも、作用する点まで同じとは限らない ということです。ヘキサラファンの特徴は、単に「わさび由来」であることではなく、分子構造の差が機能の差につながっているかもしれない点にあります。
この研究の意義:なぜ「二重経路」が重要なのか
この研究の大きな意義は、ヘキサラファンを 単なるKeap1阻害型のNrf2活性化成分としてではなく、二重経路型のNrf2安定化成分として捉え直す視点 を与えたことです。1
加齢や慢性疾患では、Nrf2が落ち込みやすい一方で、Gsk-3βの活性が高まりやすい状況が報告されています。もしその両方が同時に起きているなら、Keap1だけを標的にするよりも、Keap1とGsk-3βの両方に触れられる成分 のほうが、理論上は意味を持ちやすくなります。3 4
たとえばアルツハイマー型認知症では、細胞に酸化ストレスや炎症が起きています。それに加えて、暴走したGsk-3βが、神経細胞にたまる「タウ」という異常なタンパク質にも関わることが分かっています(ヘキサラファンとタウ病理の関係についてはこちらの記事で詳しく解説しています)。 2型糖尿病でも事情は似ています。血糖を調整するインスリンの信号がうまく働かなくなり、そこに酸化ストレスも重なります。この2つがぶつかる場所に、ちょうどGsk-3βとNrf2が居合わせているのです。 どちらの病気でも、「Nrf2が弱る」ことと「Gsk-3βが暴れる」ことが同時に起きています。だからこそ、その両方に働きかけられるヘキサラファンが、複数の病気の場面で注目される――今回の研究は、その理由を分子のレベルで説明してくれた一報だといえます。4
限界と今後の課題
一方で、この研究にははっきりした限界もあります。
まず、中心になっているのは 培養細胞と計算科学のデータ です。したがって、この論文だけで「ヒトでこう働く」とまでは言えません。実際の体内では、吸収・代謝・臓器への到達性など、さらに多くの条件が関わります。動物などの生体内(in vivo)での検証が今後必要です。1
次に、ATPポケットを標的とする分子は、理論上 ほかの酵素にも作用する可能性 があります。Gsk-3βに対して見えた作用がどこまで選択的なのかは、今後さらに検証が必要です。こうした意図しない別の標的への作用は、オフターゲット※とも呼ばれます。
用語解説- オフターゲット
- 本来狙った分子以外にも、薬剤や成分が作用してしまうこと。
さらに、Nrf2は一般に防御的ですが、高ければ高いほどよい と単純に言えるわけではありません。どの病態で、どの程度、どの期間Nrf2を安定化させることが望ましいのかという視点が、今後重要になると考えられます。3
まとめ
- ヘキサラファン(6-MSITC)は、従来知られていた Keap1阻害 に加え、Gsk-3βの直接阻害 を通じてもNrf2を安定化させる可能性が示されました。
- Keap1欠損細胞でもNrf2の増加が残ったこと、代表的な間接シグナルの変化が乏しかったこと、Gsk-3β関連の実験がそろっていたことから、Keap1に依存しない第二経路 がかなり具体的に見えてきました。
- スルフォラファンとの違いには、炭素鎖の長さによる結合の安定性の差 が関わる可能性があります。
- 加齢や慢性疾患で Nrf2低下 × Gsk-3β亢進 が重なる病態を考えるうえで、本研究はヘキサラファンの位置づけを一段深める知見といえます。
- ただし現段階では細胞・計算科学が中心であり、in vivoやヒトでの検証は今後の課題 です。
[ 引用・参考文献 ]
- García-Yagüe AJ, Cueto-Díaz EJ, Escoll M, Okunishi I, Hayes JD, Rodríguez-Franco MI, Rojo AI, Cuadrado A. Dual targeting of Keap1 and Gsk-3 by hexaraphane in the regulation of transcription factor Nrf2. Free Radical Biology and Medicine. 2025;239:579-593. ↩︎
- Rada P, Rojo AI, Chowdhry S, McMahon M, Hayes JD, Cuadrado A. SCF/β-TrCP promotes glycogen synthase kinase 3-dependent degradation of the Nrf2 transcription factor in a Keap1-independent manner. Molecular and Cellular Biology. 2011;31(6):1121-1133. ↩︎
- Cuadrado A, Rojo AI, Wells G, et al. Therapeutic targeting of the NRF2 and KEAP1 partnership in chronic diseases. Nature Reviews Drug Discovery. 2019;18(4):295-317. ↩︎
- Bartkowiak-Wieczorek J, Jodynis-Liebert J, Kujawska M. Methylsulfinyl Hexyl Isothiocyanate (6-MSITC) from Wasabi Is a Promising Candidate for the Treatment of Cancer, Alzheimer’s Disease, and Obesity. Nutrients. 2024;16(15):2509. ↩︎
- Yoshida J, Nomura S, Nishiumi S, et al. Glycogen synthase kinase-3beta inhibition by 6-(methylsulfinyl)hexyl isothiocyanate derived from wasabi (Wasabia japonica Matsum). Bioscience, Biotechnology, and Biochemistry. 2011;75(1):136-139. ↩︎